|
|
by Darel Rex Finley
Revised September 14, 2000 (reference to truncation selection added)
Suppose I am a credentialed scientist, with a strong non-scientific motive
to promote a theory, T. My motivation might be philosophical, or I might have
invested so much of my career in theory T that it would be personally disastrous
to relinquish it. Whatever the case, I privately consider T to be an unquestionable
Truth, but in order to convince the population at large, I must pay lip
service to the status of T as a scientific theory.
I begin by pointing to the existing body of supporting evidence, and declare
T to be a fully confirmed (and therefore true) theory.
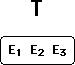
E1, E2 and E3 are items of empirical evidence that support theory T.
Now, along comes a new piece of not-so-friendly evidence, E4.
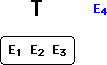
If T and E4 are compared in isolation, E4 does not work very well with
T, perhaps even disconfirms it. At the very least, T did not predict that
E4 would ever be observed.
So to protect T, I employ the following logic: E1-3 prove that T must be
true. The truth of T means that observation E4 demands an explanation, X4,
which attempts to reconcile E4 with the theory T.
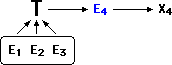
Explanation X4 may be completely ad hoc, unsubstantiated, far-fetched
etc., but it (or something like it) must be true, because the truth
of T demands it.
Next, I simply reverse the order of E4 and X4.
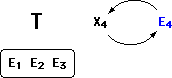
Now, the theory T, refined by explanation X4, predicts evidence E4. Since
E4 was in fact empirically observed, the theory is confirmed by E4!
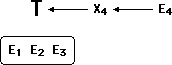
E4 can now be drawn into the fold of supporting evidence. (Explanation
X4 is quietly dropped from the picture, to be revived any time the confirmatory
status of E4 is challenged.)
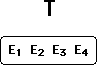
Then, the process can be repeated. A new piece of unfriendly evidence
E5 is found. Explanation X5 is demanded by the manifest truth of T (as evidenced
by E1-4).
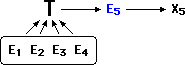
X5 and E5 are reversed, and E5 is now what T would predict, hence confirmatory.
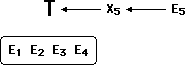
By this method, the theory can be marvelously protected from any number of hostile discoveries. As time goes by, the body of "confirming" evidence grows larger and larger, and the soundness of this process seems more and more irrefutable.
There are four problems with this defense tactic, however.
1. Non-absolute Proof The whole logic of the process depends on
the original body of supporting evidence (E1, E2, E3) constituting absolute
proof of T, beyond any possible doubt. If E1-3 leave any doubt,
then it is possible that new evidence, such as E4, E5, E6, might eventually
substantiate such doubts. But such substantiation can never be revealed
if any disconfirming qualities of E4+ are obfuscated in the name of E1-3.
2. Tautology The reversal of E4 and X4 is a circular argument, and
is the source of the frequently tautological nature of this kind of defense. X4 is demanded by E4, then E4 is predicted by X4! Tautologies do not verify
scientific theories.
3. Lost Explanations Lost in this process are the ad hoc
explanations, X4, X5 etc. Rationally, they should be incorporated into
theory T, where they might need their own confirmation, be subject to possible
falsification, and be compared with one another for consistency. Instead,
they are simply stashed away in a hidden place, to be pulled out and cited
when convenient. The original theory T is left in pristine condition, and
the "confirmatory" status of E4+ is applied to that unmodified
version of T. This amounts to a sophisticated form of intellectual dishonesty.
4. Unscientific Method Admittedly, the scientific method does not
preclude the declaration of a theory as amost-certainly confirmed. Neither
does it preclude the presumption that a heavily confirmed theory is true,
when exploring other areas of the scientific frontier. But it does preclude
the interpretation of new evidence under the presumption that a theory
can never be falsified. The tactic of theory defense outlined above
depends entirely on just such a violation of the scientific method.
When the anomalous evidence of sudden appearance and stasis in the fossil record became apparent, the ad hoc explanation of "punctuated equilibrium" was devised. This explanation has not been treated as a hypothetical in need of confirmation, but is simply cited when critics bring up the issue of sudden appearance and stasis.
Allen Orr's "Part A, Part B" response, to the issue of irreducible complexity, is also extremely ad hoc, and should properly require numerous molecular supporting examples (and no refuting ones) before it can be considered a re-establishment of the scientific validity of evolution by natural selection. So far, however, the scientific community seems contented to let Orr use his scenario against Behe, while leaving evolution, as generally taught, unmodified by Orr's scenario.
As Walter ReMine documents in The Biotic Message, Haldane's dillemma (available time) was never solved, and "truncation selection" is invoked as the hypothetical solution when the subject comes up. But the subject rarely comes up, and so standard evolutionary genetics is taught unammended by truncation selection or any other purported solution.
To most evolutionary scientists, the idea that after all these years, religious creationists might be right — even partially right — is unthinkable. But science is not religion-bashing. Science is an objective, rational search for empirically verifiable truth, without concern for which historical figures may be vindicated, nor who may benefit. If the National Academy of Sciences announced that Darwin's evolution had crumbled under the weight of new evidence, and that intelligent design was a viable possibility, biblical literalists like Duane Gish would cheer while scientists like Richard Dawkins and Francis Crick would cry foul, but the announcement would be no less scientific for it.

After I posted Why I Disbelieve Evolution on the Web, I received e-mail from a lot of people, some of them more rational than others. I have distilled their best arguments into a set of ten questions, and attempt to answer each as best I can.
1. Even if Darwin's theory has been suffering a host of empirical problems lately, we can be confident that a naturalistic explanation will be found, because even if Darwin's theory is wrong, he at least proved that a naturalistic explanation of life is possible, right?
This is a close parallel to Paul Davies's argument that Stephen Hawking's "no boundary" proposal proves that the universe wasn't created, even if the proposal is false. As described by Johnson on page 58 of Reason In the Balance:
|
Paul Davies, another prominent mathematical physicist, explains the importance of Hawking's no-boundary proposal in his book The Mind of God, in a chapter titled "Can the Universe Create Itself?" Davies candidly states at the outset, "This particular explanation may be quite wrong," but immediately adds that the correctness of any particular explanation does not matter. According to Davies, "What is at issue is whether or not some sort of supernatural act is necessary to start the universe off. If a plausible scientific theory can be constructed that will explain the origin of the entire physical universe, then at least we know a scientific explanation is possible, whether or not the current theory is right."
|
The answer to Davies, and to anyone who would apply the same logic to evolution, is: No — explanations that turn out to be false do not prove anything.
This is not a trivial issue; it goes to the heart of the E4->X4 method of theory defense outlined above. The basic idea is that if evolution seemed credible under the evidence that was known in Darwin's time, that somehow makes it "scientific" to defend it today. But it really doesn't. There's a lot of truth in the idea that "ignorance is innocence," but the pretense of ignorance is far from innocent. Darwin can be forgiven for thinking that natural selection could create the functions of life, for he was ignorant of how those functions worked. Modern evolutionists cannot be excused for pretending not to know better than Darwin did in 1859.
2. A lot of the scientists quoted in books by Johnson, Denton and Behe are firm believers in evolution. Aren't they being quoted out of context, and doesn't that invalidate those anti-evolution arguments?
"Quoting out of context" implies that the meaning of the quotation is being distorted by the removal of the context, since the context was necessary to understand the meaning of the quotation. But the intended conclusion of the author, or his public statements of support for evolution, are not "context" required for the correct understanding of specific quotations. For example, if Michael Denton quotes Francis Crick as having said
|
An honest man, armed with the knowledge available to us now, could only state that in some sense, the origin of life appears at the moment to be almost a miracle, so many are the conditions which would have to have been satisfied to get it going. —Crick, as quoted in Evolution: A Theory In Crisis, page 268
|
and uses that quotation to bolster the idea that the origin of life perhaps was a miracle, this does not constitute a distortion of the meaning of that quotation, even though Crick — as an openly atheist evolutionist — might personally dislike its use by Denton.
3. Phillip Johnson admits that he is a theist and Christian. Isn't this an ulterior motive? How can we trust his empirical arguments when he has an obviously non-empirical ulterior motive?
Suppose, upon opening Darwin On Trial, we found the words
|
Evolution is false — trust me.
The End
|
and the rest of the pages were blank. In that case, the above argument would apply. It does not apply to the book Johnson actually wrote, because Johnson is not asking us to trust him — that's why he filled his book with empirical arguments.
What the "trust" attack seems to be suggesting is that Johnson knows of empirical refutations of his own thesis, but keeps them secret in order to advance biblical literalism. Does only Johnson know of these refutations? Is Phillip Johnson, Professor of Law, the lone carrier of the secret empirical proof of evolution? Clearly this is ridiculous. If evolutionists want us to believe that Professor Johnson is advancing a bad argument, they need to tell us what is wrong with the argument. If they cannot, then they are merely substituting ad hominem attacks for real science.
(None of this should be taken to imply that discussion of ulterior motives is off-limits; rather that it should accompany substantive refutation, not substitute for it.)
4. Defining "fitness" to mean "survival" has been criticized, since it turns the phrase "survival of the fittest" into a tautology. But tautology or not, doesn't it still prove that Darwin's thesis is true?
Not hardly. Consider the following diagram:
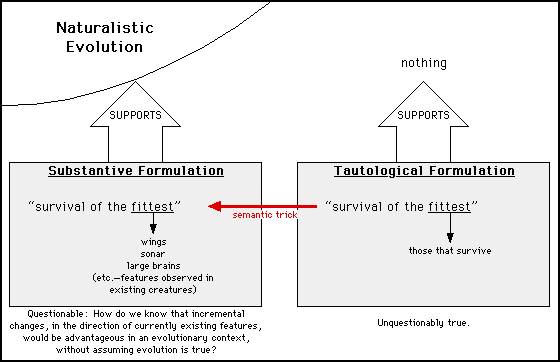
When discussing evolution in an uncritical forum, proponents draw upon the substantive formulation of "survival of the fittest," to support the idea of biological evolution. When this formulation comes under attack, they switch to the tautological formulation, which is immune from criticism but does not support evolution or anything else.
The red arrow in the middle — representing the fact that the two phrases are composed of the same words — is the bridge between "Unquestionably true" and "Naturalistic Evolution." The logical flaw is that redefining the word "fittest" changes the meaning of the phrase, and there is no logical link between two concepts of different meaning simply because they are frequently identified with the same sequence of words.
This semantic trick is sometimes employed with other evolutionary phrases:
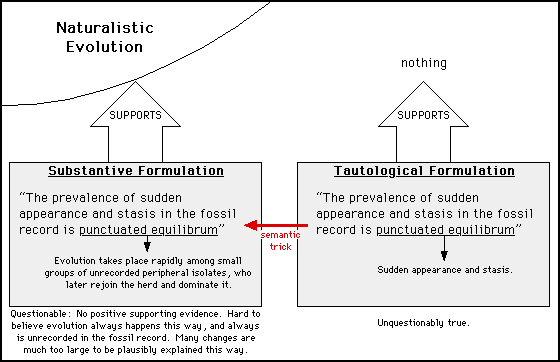
In this case, the phrase "punctuated equilibrium" is being manipulated. Tony Schountz of the Oak Ridge National Laboratory in Tennessee used this stunt in his e-mail to me, when he said that by describing sudden appearance and stasis I was simply describing "punctuated equilibrium" which supports evolution. To say I was describing punctuated equilibrium is to employ the tautological formulation, which does not support evolution. "Punctuated equilibrium," in its substantive form, is a very hypothetical explanation of sudden appearance and stasis — not a restatement of those phenomena.
"Fitness" and "punctuated equilibrium" can be and are manipulated, but the most highly manipulated word in the evolutionary vocabulary is "evolution" itself. Here is just one example:
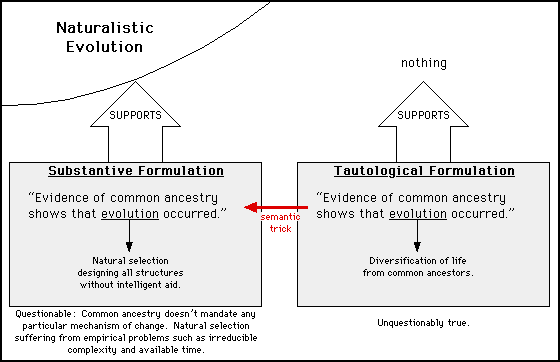
In Darwin's Black Box, Michael Behe said that he believes common ancestry to be "fairly convincing," but he had much criticism of evolution. When Allen Orr accused Behe of hypocrisy for holding these allegedly contradictory positions, Orr was employing the tautological formulation, which equates evolution to common ancestry by definition. But Behe was expressing his disbelief in the substantive definition of evolution, and thus was not being hypocritical.
Johnson eloquently described the situation on page 151 of Darwin On Trial (page 153 of the paperback edition):
|
If critics are sophisticated enough to see that population variations have nothing to do with major transformations, Darwinists can disavow the argument from microevolution and point to relationship as the "fact of evolution." Or, they can turn to biogeography, and point out that species on offshore islands closely resemble those on the nearby mainland. Because "evolution" means so many different things, almost any example will do. The trick is always to prove one of the modest meanings of the term, and treat it as proof of the complete metaphysical system.
|
5. The fossil record may suffer from sudden appearance and stasis, but doesn't it still present a general picture of increasing sophistication that is strongly indicative of evolution?
The problem of sudden appearance and stasis cannot be brushed aside simply because doing so might make evolution whole. Many evolutionary texts do exactly that; they show diagrams in which fossil species are connected together through time by broad, branching lines of evolution. The implication of these connecting lines is that the diagram has been simplified for the sake of brevity; to show all the transitionals would crowd the page. Often, however, the diagram is not simplified at all — the transitionals are not shown on paper because they are not shown in the fossil record, and the lines connecting them are imaginary.
The following five graphs are heavily simplified, but I think the points they make are still valid:

Graph 1: Implication of Evolution
The above graph illustrates what evolution should look like. Not only must transitions be continuous, but the rate of evolution must rapidly decelerate. This is for three reasons.
Generational cycle times: Simpler, smaller creatures multiply much faster. A bacterium can divide many times in a day, while the typical human baby is born to parents perhaps about twenty years old. Since evolution is driven by differential survival of variant offspring, shorter generation cycles should mean faster evolution.
Numbers of offspring: Simpler, smaller creatures have many more offspring. Insects lay thousands of eggs at a time, while humans typically have only one baby at a time. A large number of offspring dramatically increases the chance that one will be conferred with an advantageous mutation.
Survival rates: The survival rate for large sophisticated creatures, cared for and nurtured by parents, is generally much higher than the survival rate for say, fish, whose young grow up on their own in a highly predatory environment with nowhere near the resources necessary for more than a tiny percentage to survive. Hence, only the very strongest and most adapted fish will pass on their genes to the next generation. Evolution should be more efficient under these conditions.
The following graph illustrates the problem:
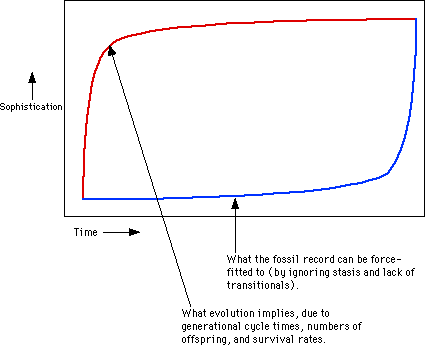
Graph 2: Evolution Rates
The red line has high evolution rates for simpler life forms, and a dramatic slowdown as creatures become more sophisticated. But the blue line is more like what the fossil record really shows. Could these lines be more different?
The fossil record actually looks something like this:

Graph 3: The Fossil Record
The leap from primates to man could be reasonably considered much taller than depicted here. The human brain alone represents a leap more fantastic than any other in the history of life, and humans are more sophisticated than primates in other ways as well.
Explaining how evolution by natural selection acting on mutation could have produced a fossil record that looks like this is a vastly bigger problem than finding a few passable intermediates like Archaeopteryx.
It should be noted that Graph 1 (Implication of Evolution) is actually being rather kind to the theory. Notice that although the evolutionary rates decelerate as evolution implies they should, the rates are still not consistent with the levels of sophistication. There just isn't any way to do that and still have a common ancestor. A graph with consistent rates would look like this:
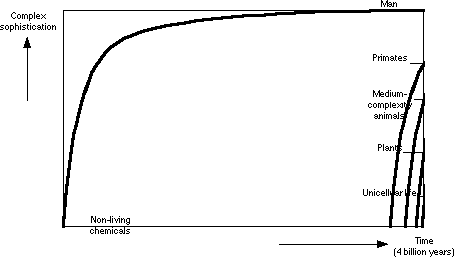
Graph 4: Consistent Rates of Evolution
Now the rates are consistent, but at what cost? Common ancestry is destroyed and replaced with multiple origins of life, which become inexplicably more frequent near the present. Unicellular life is absent for large stretches after its first appearance. Obviously, this just won't do. And no evolutionists are promoting it.
What they are promoting, though, is this:
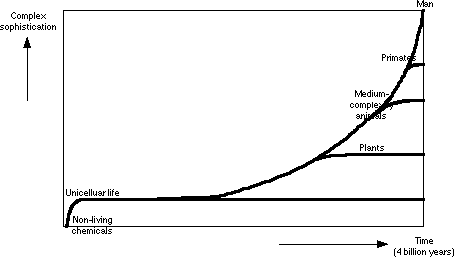
Graph 5: Evolutionary Orthodoxy
The Evolutionary Orthodoxy graph contradicts the implications of the theory regarding evolutionary rates, and only superficially resembles the fossil record. So why are the fossils presented to the public as a huge, undeniable proof of evolution? Because the theory has to be protected.
6. You calculated that the average rate of evolution would need to be a highly unrealistic 0.75 base pairs per year. Since the human genome is thought to be up to 95% junk (unused, duplicates, etc.), doesn't this save the theory?
I don't know how the estimate of "up to 95% junk" is derived, but let's assume it's true. Then, the average rate could be as slow as one base pair every 26.7 years. Given that primates have generation cycles of at least a few years, even if a beneficial mutation spread with 100% efficiency (passed on to every descendent without exception) it would still take maybe 26.7 years for it to spread through a population of thousands. This means that beneficial mutations writing the present code of the human genome must have been arriving every quarter-century or so among pre-human primates. The most evolutionally optimistic estimates of the frequency of beneficial mutation rates can't conceivably come close to the required rate!
If the beneficial mutation is only slightly beneficial, then the efficiency with which it spreads through the population would be very low. It could take millennia for the mutation to dominate the population, and there would be a fair chance that the near-random walk of inheritance would kill the mutation shortly after its appearance, by bad luck.
The use of primates as an example can be objected to, since one-per-26.7 years is the average rate, and the rate might be much faster for lower species. But a realistic estimate of beneficial mutation rates in lower species is still likely to be abysmally short of the requirement. And the advancements of humans above primates are large, not trivial. Also, mutation rates (beneficial and detrimental combined) must be limited to what the species can survive.
That is why I said in my original article that the time available might need to be in the trillions or quadrillions of years. Maybe if the human genome were found to be 99.9999% junk, I would start wondering if the Available Time problem had been solved. The other problems would remain. What I called "feature interdependence," and Michael Behe has christened "irreducible complexity," indicates that a staircase of incrementally beneficial mutations leading to the human genome does not exist, and therefore could not be ascended in 10100 years.
7. Mutations can easily cause large-scale changes in the capabilities of individuals, such as a mutation that causes blindness by jamming the light-sensing system of retinal nerve cells. Then, natural selection quickly acts on the differential survival rates and selects for the sighted individuals. Doesn't this conclusively prove Darwinian evolution?
This same argument was used by Brown University biology professor Kenneth Miller in a NOVA Internet debate with Phillip Johnson. Miller wrote:
|
Indeed, the most recent issue of Science (Nov-15, page 1082) reported that a single gene controls tunicate tail formation. Mutate it, the tail is lost. Restore it, tail comes back. Just another example of a genetic mechanism producing macroevolutionary change.
|
(Note that a "gene" is a rather large quantity of DNA code.)
The fallacy of this argument is that it confuses destructive evolution with constructive evolution. Just as it is extremely easy for a tornado to destroy a house, but could never be expected to build one, so it is very easy for a random mutation to jam a molecular machine, but not to create one! Even a large number of mutations spread out over a very long time could prove unable to create irreducibly complex systems.
The ability of natural selection to efficiently preserve a sophisticated mechanical function once it exists, does little to explain what made the function in the first place.
8. Didn't the Grants prove evolution by their studies of finch beaks? The beaks rapidly changed in size as a response to changes in the food supply.
This question is actually very similar to question 7.
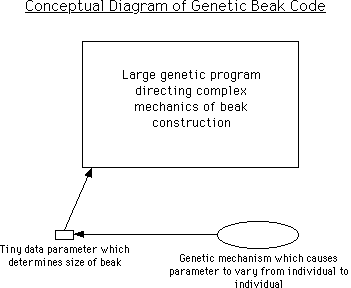
What the above diagram illustrates is that a beak can easily change size when its size is controlled by a tiny data parameter, which also is set up to be highly variable from one generation to the next. This situation does little or nothing to explain where the large — and perhaps irreducibly complex — beak construction program came from.
9. Darwin said "If it could be proved that any part of the structure of any one species had been formed for the exclusive good of another species, it would annihilate my theory, for such could not have been produced through natural selection." As Darwin predicted, nature seems devoid of such features. For example, bees benefit flowers by pollinating them, but the bees also benefit by the nectar they collect from the flowers. Why wouldn't a designer just program the bees to spend some of their time pollinating flowers, with no benefit to the bee?
A designer might find symbiosis esthetically pleasing. Or, the goal of efficiency might be served by having the bees pollinate flowers and collect food at the same time, instead of as separate tasks.
But the most important reason is very close to the one Darwin believed. If a designer gave the bee a genetic program causing it to instinctively pollinate flowers at no benefit to the hive, then what would happen when a chance mutation jammed the program, and a queen was formed that produced non-pollinating drones? Her hive would be more efficient, since her drones would waste no time in non-beneficial tasks. Instead of eliminating the mutation, natural selection would favor the defective bees, and they would wipe out the normal bees. Then, the bee population would still contain the genetic program for pollination, but in a broken, deactivated form, and the flowers would not get pollinated.
(Would that scenario be an example of evolution? Yes it would, but it would be an example of destructive evolution — see question 7.)
It also works the other way: Suppose that the designer programmed the flower to produce nectar for bees, at no benefit to the flower. Then, if a chance mutation jammed the nectar production program, natural selection would not weed out that mutation, and might actually favor it if the flower was more efficient than its nectar-producing brethren.
So, unless the designer wanted to be repeatedly intervening to fix his work, the symbiotic arrangement would be preferable.
10. When environmental conditions change dramatically, this pressures creatures to evolve, doesn't it? Environmental pressure is the mechanism of evolution.
"Environmental pressure," also called "selection pressure," is an undefined force which allegedly pushes creatures into new forms. I reject this idea, because if the environment changes drastically, this may create a need for new adaptations, but does not provide an empirically sound mechanism for creating those adaptations. Calling "environmental pressure" a "mechanism" of evolution is to employ the semantic trick discussed in question 4.
When environmental shifts necessitate new adaptations, the species does not have to acquire them; it has another perfectly acceptable alternative: Become extinct. And if there is no mechanism — natural or otherwise — for a species to adapt to severe environmental changes, then become extinct it will.
The July 25 1997 issue of Science contains a paper titled "Evidence For A Large-Scale Reorganization of Early Cambrian Continental Masses By Inertial Interchange True Polar Wander" by Kirschvink, Ripperdan, and Evans. The article is a highly technical proposal that Earth underwent extremely rapid and unusual continental shifts around the same time as the "Cambrian explosion" — the time when hundreds of species representing all animal phyla appeared suddenly and without evolutionary history.
Evolution is mentioned twice in this lengthy paper — first at the beginning of the article:
|
The age constraints on the geophysical data indicate that the rapid continental motions occurred during the same time interval as the Cambrian evolutionary diversification and therefore the two events may be related.
|
and again at the end of the article:
|
The repeated reorganizations in global climatic patterns during an IITPW event should have fragmented any large-scale ecosystems that were established, generating smaller, more isolated populations and leading to a higher evolutionary branching rate among existing groups. Comparisons of evolutionary rates between Cambrian and late Paleozoic time, corrected for the new time scale, indicates that the Cambrian rate was a factor of ~20 times higher.
|
Although the article did not specifically say that the continental shifts prove the theory of evolution by natural selection, the authors did nothing to dissuade such an interpretation, and so their research was reported by CNN this way:
|
One-half billion years ago, for reasons scientists have never clearly understood, there was an unprecedented burst of evolutionary activity, greater than any before or since.
. . .
If [the continental shift] took place, cold continents would have thawed and tropical continents would have cooled. Similar upheavals would have taken place in the oceans. And the ensuing environmental changes caused by all of this created the chaotic conditions needed for rapid evolution. (July 25, 1997)
|
and FOX News portrayed it so:
|
Scientists have long wondered what caused the sudden evolutionary spurt known as the "big bang." In a report published today in the journal Science a group of geologists present what may be the answer.
The "big bang" began about 530 million years ago when evolution proceeded at a rate 20 times faster than anything that has happened since. Scientists now believe that a 90-degree shift of the Earth's early continents . . . may have played a major role in this evolutionary explosion and speeded up the development of life. (July 25, 1997)
|
These are perfect examples of the "environmental pressure" fallacy. If the proposed continental shifts did occur, and did so at about the same time as the Cambrian explosion, that's an interesting correlation, but it still leaves unanswered the question: By what biological mechanism did evolution proceed from single celled organisms to large, complex multicellular organisms, so quickly that there is no fossil documentation of the transition?
|


















